Inhalt der Folge:
- In dieser Podcastfolge besprechen wir die Elongationsphase der DNA Replikation.
Die Phasen der DNA Replikation
Der Ablauf der DNA Replikation lässt sich in drei Phasen unterteilen:
- Die Initiationsphase (Folge 035)
- Die Elongationsphase (Thema dieser Folge)
- Die Terminationsphase
Die Elongationsphase
Die Elongationsphase der DNA Replikation folgt direkt im Anschluss an die Initiationsphase und kann ebenfalls in mehrere Reaktionsschritte geliedert werden:
Schritt 1: Priming
- Im Anschluss an die Öffnung des DNA-Doppelstranges in der Initiationsphase folgt das sogenannte Priming.
- Unter dem Priming versteht man die Synthese der Primer an den DNA-Einzelsträngen.
- Primer sind kurze Stücke RNA, die ca. 6-30 Nukleotide lang sind.
- Die Synthese der Primer erfolgt durch das Enzym Primase (eine bestimmte RNA-Polymerase).
- Die Primase benötigt lediglich einen DNA-Einzelstrang als Vorlage, um die Primer zu synthetisieren.
- Sie liest den DNA-Einzelstrang, der als Vorlage dient, in 3′ → 5′ Richtung ab und synthetisiert einen komplementären Primer in 5′ → 3′ Richtung.
Die Funktion der Primer
- Dass die DNA erst nach dem Priming repliziert werden kann, liegt an dem Enzym, das die neuen DNA-Einzelstränge synthetisiert.
- Dieses Enzym heißt DNA-Polymerase (genauer gesagt DNA-Polymerase 3).
- Die DNA-Polymerase braucht immer eine freie 3′-OH-Gruppe, um mit der Synthese eines neuen DNA-Einzelstranges beginnen zu können.
- Diese freie 3′-OH-Gruppe wird durch die Primer bereitgestellt.
- Die Primer dienen der DNA-Polymerase also als Starthilfe.
Die DNA Polymerase
- Aufgabe der DNA-Polymerase ist es, die neuen DNA-Einzelstränge zu synthetisieren.
- Sie liest die Basen der DNA-Einzelstränge, die als Kopiervorlage dienen, nacheinander ab und synthetisiert Base um Base einen komplementären Strang nach dem Prinzip der komplementären Basenpaarung (A&T / C&G).
- Wichtig zu verstehen ist die 5’→3′-Aktivität der DNA-Polymerase, mit der die Richtung der Synthese gemeint ist.
- Die DNA-Polymerase liest den DNA-Einzelstrang, der als Kopiervorlage dient, immer in 3′ → 5′ Richtung ab und kann den neuen Strang nur in 5′ → 3′ Richtung synthetisieren!
- Sie stellt die einzelnen Nukleotide jedoch nicht selbst her, sondern verknüpft sie lediglich.
- Die benötigten Nukleotide befinden sich bereits zu genüge im Cytoplasma der Zellen.
- Für die Synthese von Nukleotiden ist wiederum größtenteils die Leber verantwortlich.
Die Problematik der Antiparallelität: Leitstrang und Folgestrang
- Die beiden Einzelstränge einer Doppelsträngigen-DNA verlaufen antiparallel.
- Das bedeutet, dass die Einzelstränge zwar parallel verlaufen, jedoch entgegengesetzt gerichtet sind.
- Mit anderen Worten: Dort wo der eine DNA-Einzelstrang sein 3′-Ende hat, hat der andere Strang sein 5′-Ende und umgekehrt:
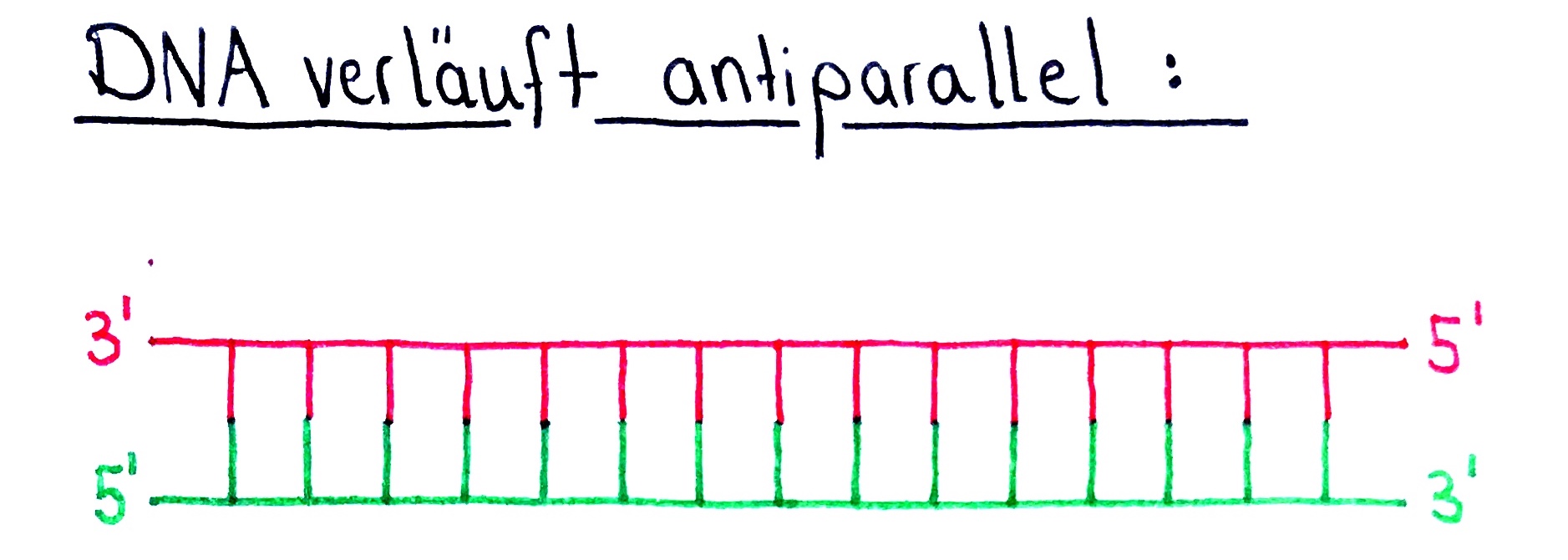
- Die DNA-Polymerase kann aber nur in 5′ → 3′ Richtung synthetisieren bzw. den Elternstrang in 3′ → 5′ Richtung ablesen.
- Mit voranschreitender Öffnung der Replikationsgabel kann also nur einer der beiden Stränge in Öffnungsrichtung (3′ → 5′ Richtung) abgelesen werden.
- Dieser Strang wird Leitstrang genannt.
- Der andere Elternstrang wird Folgestrang genannt.
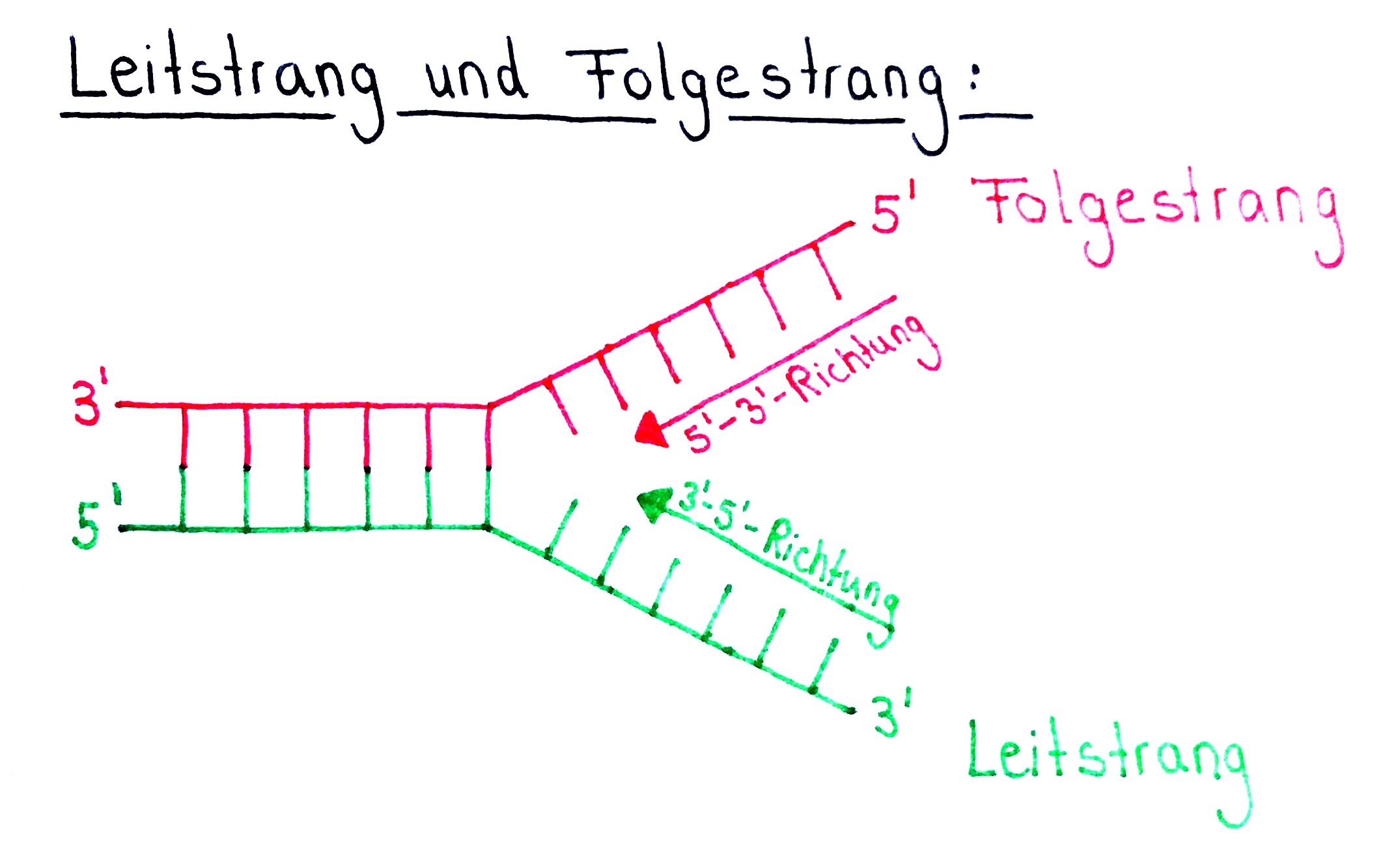
Es gibt nicht nur einen Leitstrang und einen Folgestrang!
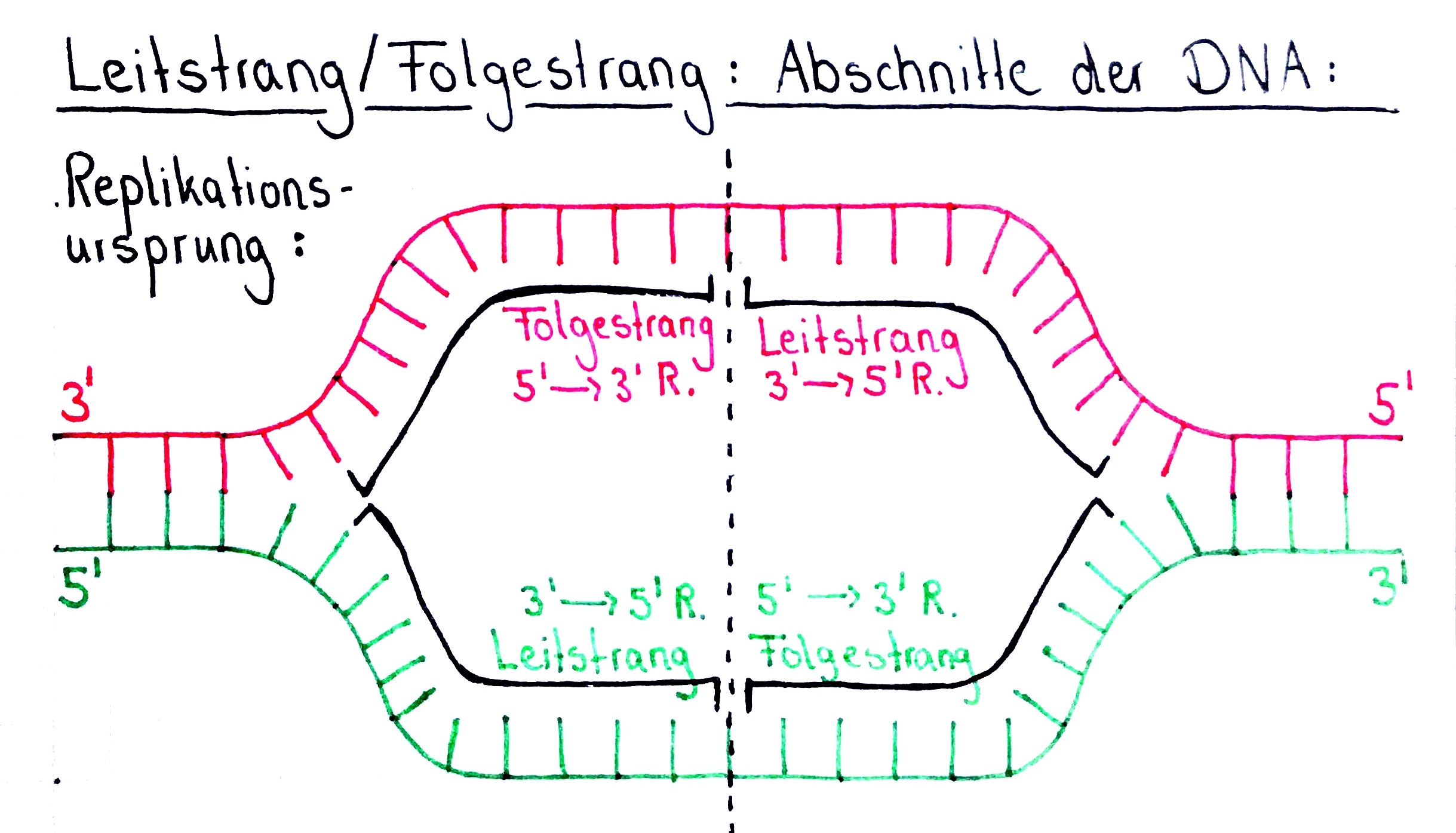
- Wichtig zu verstehen ist, dass die Bezeichnungen „Leitstrang“ und „Folgestrang“ immer nur im Hinblick auf eine einzelne Replikationsgabel Sinn ergeben.
- Wie wir bereits aus der letzten Folge wissen, wird die Eltern-DNA während der Initiationsphase an den Replikationsursprüngen geöffnet.
- Man muss sich jedoch klar machen, dass im Anschluss zwei Replikationsgabeln entstehen.
- Denn ausgehend vom Replikationsursprung wird die Eltern DNA in beide Richtungen von Helicasen geöffnet.
- Deshalb kann es nicht nur einen Leitstrang und einen Folgestrang geben, wie wir anhand dieser Abbildung noch besser nachvollziehen können:
Schritt 2: DNA-Synthese am Leitstrang
- Die DNA-Synthese am Leitstrang ist schnell erklärt und relativ unkompliziert.
- Der Leitstrang kann in Öffnungsrichtung durchgehend repliziert werden, da hier die Öffnungsrichtung der Leserichtung der DNA-Polymerase (3′ → 5′ Richtung) entspricht.
- Nach einmaligem Priming gleitet die DNA-Polymerase am Leitstrang in 3′ → 5′ Richtung entlang und synthetisiert kontinuierlich den neuen Strang in 5′ → 3′ Richtung.
Schritt 3: DNA-Synthese am Folgestrang
- Die DNA-Synthese am Folgestrang ist ebenfalls schnell verstanden, verläuft aber etwas komplizierter.
- Am Folgestrang ist nämlich keine kontinuierliche Replikation möglich, da hier die Öffnungsrichtung nicht der Leserichtung der DNA-Polymerase entspricht.
- Denn wenn man in Öffnungsrichtung schaut, verläuft der Folgestrang in 5′ → 3′ Richtung.
- Die Leserichtung der DNA-Polymerase ist jedoch nicht von 5′ → 3′, sondern von 3′ → 5′.
- Die Replikation am Folgestrang kann also erst ablaufen, wenn die Eltern-DNA bereits ein Stück geöffnet ist.
- Im Anschluss erfolgt das Priming direkt hinter der Helicase.
- Danach wird der neue DNA-Strang gegen die Öffnungsrichtung diskontinuierlich synthetisiert.
Okazaki-Fragmente
Das erste Priming
- Nach dem ersten Priming am Folgestrang synthetisiert die DNA-Polymerase den neuen Strang gegen die Öffnungsrichtung.
- Nach kurzer Zeit trifft die DNA-Polymerase allerdings auf den Primer des Leitstranges der Replikationsgabel, die in die andere Richtung verläuft.
- Sobald die DNA-Polymerase auf diesen Primer trifft, wird die DNA-Synthese an dieser Stelle abgebrochen.
Das zweite Priming
- Währenddessen öffnet die Helicase die Eltern-DNA munter weiter.
- Um den nächsten Abschnitt des Folgestranges replizieren zu können, muss es erneut zum Priming kommen, um der DNA-Polymerase wieder „Starthilfe“ zu geben.
- Nach diesem zweiten Priming am Folgestrang synthetisiert die DNA-Polymerase den neuen Strang wieder gegen die Öffnungsrichtung.
- Nach kurzer Zeit trifft die DNA-Polymerase logischerweise auf den ersten Primer des Folgestranges und bricht die DNA-Synthese an dieser Stelle wieder ab.
- Dieser Ablauf wiederholt sich so oft, bis der gesamte Folgestrang repliziert wurde.
- Dabei entstehen viele einzelne Abschnitte, bestehend aus neu synthetisierter DNA (immer ca. 1000 – 2000 Nukleotide lang) und dem jeweiligen Primer.
- Diese Abschnitte nennt man Okazaki-Fragmente.
Schritt 4: Die Beseitigung der Primer der Okazaki-Fragmente
- Um einen durchgehenden DNA-Strang zu bekommen, der keine RNA-Stücke (Primer) mehr enthält, müssen diese entfernt und durch DNA ersetzt werden.
- Eine bestimmte Ribonuklease (Enzyme die Phosphodiesterbindungen spalten können) ist in der Lage die Primer zu entfernen.
- Eine weitere DNA-Polymerase (DNA-Polymerase I) füllt die entstandenen Lücken mit der jeweils komplementären DNA.
- Dieser Vorgang findet bereits während der Replikation des Folgestranges statt und nicht erst im Anschluss.
Schritt 5: Die Ligation der Okazaki-Fragmente
- Der fünfte und letzte Schritt der Elongationsphase ist die Verknüpfung der Okazaki-Fragmente.
- Zwar wurden die Primer bereits durch DNA ersetzt, allerdings befinden sich noch kleine Lücke zwischen den 3′-Enden und den 5′-der jeweiligen Okazaki-Fragmente.
- Das Enzym Ligase kann diese letzten Lücken schließen, indem es die freien 3′-OH-Gruppen mit den freien 5′-Phosphatgruppen verknüpft.
Und damit ist die Elongationsphase der DNA Replikation beendet und wir machen in der nächsten Folge weiter mit der dritten und letzten Phase.
Die DNA Replikation ist semidiskontinuierlich
- Der Leitstrang wird ohne Unterbrechungen in Öffnungsrichtung repliziert, deshalb spricht man beim Leitstrang von einer kontinuierlichen Replikation.
- Weil der Folgestrang nicht kontinuierlich, sondern in Abschnitten synthetisiert wird, spricht man beim Folgestrang von einer diskontinuierlichen Replikation.
- Betrachtet man die Replikation als Ganzes, muss man die beiden Begriffe „kontinuierlich“ und „diskontinuierlich“ lediglich kombinieren und bekommt den Begriff „semidiskontinuierlich“.
- Deshalb bezeichnet man die DNA Replikation als semidiskontinuierlich.
